Univ.-Prof. Dr. Holger Daims

Links to CV and publication output summaries are available at the bottom of this page.
Nitrification, Comammox, and Nitrite-oxidizing bacteria
 Nitrification, the microbially catalyzed oxidation of ammonia to nitrate via nitrite, is an essential process of the biogeochemical nitrogen (N) cycle. Nitrification is the primary biological source of nitrate, which is an important N source for many microorganisms and plants and a main electron acceptor for anaerobic respiration. Being a key process of biological wastewater treatment, nitrification is highly important for protecting our environment from anthropogenic pollution. On the other hand, nitrification contributes to N losses from soils and thus it negatively affects the efficiency of fertilization in agriculture. Nitrification is carried out by ammonia-oxidizing bacteria (AOB) and archaea (AOA), complete ammonia oxidizers (comammox), and nitrite-oxidizing bacteria (NOB). My research group investigates the biology and evolution of the nitrifying microorganisms, with focus on comammox and NOB. Our aim is to contribute to a better microbiological understanding of the N cycle. Such knowledge will be essential to assess how the massive discharge of N from fertilizers and sewage affects our environment and to develop strategies for dealing with these problems.
Nitrification, the microbially catalyzed oxidation of ammonia to nitrate via nitrite, is an essential process of the biogeochemical nitrogen (N) cycle. Nitrification is the primary biological source of nitrate, which is an important N source for many microorganisms and plants and a main electron acceptor for anaerobic respiration. Being a key process of biological wastewater treatment, nitrification is highly important for protecting our environment from anthropogenic pollution. On the other hand, nitrification contributes to N losses from soils and thus it negatively affects the efficiency of fertilization in agriculture. Nitrification is carried out by ammonia-oxidizing bacteria (AOB) and archaea (AOA), complete ammonia oxidizers (comammox), and nitrite-oxidizing bacteria (NOB). My research group investigates the biology and evolution of the nitrifying microorganisms, with focus on comammox and NOB. Our aim is to contribute to a better microbiological understanding of the N cycle. Such knowledge will be essential to assess how the massive discharge of N from fertilizers and sewage affects our environment and to develop strategies for dealing with these problems.
Since most nitrifiers are yet uncultured, our studies rely on a battery of cultivation-independent molecular and isotope techniques as well as “omics” approaches. In addition, we cultivate nitrifying microbes and carry out experiments with pure cultures and enrichments of these fastidious organisms.
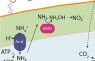
 Biology of comammox. We contributed to the discovery of complete ammonia oxidizers (comammox), which belong to the highly diverse and environmentally widespread NOB-genus Nitrospira. In contrast to all other nitrifiers, comammox are capable of complete nitrification (oxidation of ammonia to nitrate) in a single organism (Daims et al. 2015, Nature). This discovery resolved a century-old question about the N cycle and fundamentally changed our picture of nitrification. It also raised a large number of new questions ranging from the environmental distribution, ecological niche and evolution of comammox to the kinetics, regulation and biochemistry of complete nitrification. Intriguingly, comammox Nitrospira are well adapted to life in oligotrophic habitats: based on their kinetic properties, comammox can outcompete other ammonia oxidizers (including many ammonia-oxidizing archaea) under strong ammonia limitation (Kits et al. 2017, Nature). We study the biology of these organisms with our primary comammox culture (Nitrospira inopinata), new comammox enrichments, and cultivation-independent approaches in collaboration with the group of Michael Wagner.
Biology of comammox. We contributed to the discovery of complete ammonia oxidizers (comammox), which belong to the highly diverse and environmentally widespread NOB-genus Nitrospira. In contrast to all other nitrifiers, comammox are capable of complete nitrification (oxidation of ammonia to nitrate) in a single organism (Daims et al. 2015, Nature). This discovery resolved a century-old question about the N cycle and fundamentally changed our picture of nitrification. It also raised a large number of new questions ranging from the environmental distribution, ecological niche and evolution of comammox to the kinetics, regulation and biochemistry of complete nitrification. Intriguingly, comammox Nitrospira are well adapted to life in oligotrophic habitats: based on their kinetic properties, comammox can outcompete other ammonia oxidizers (including many ammonia-oxidizing archaea) under strong ammonia limitation (Kits et al. 2017, Nature). We study the biology of these organisms with our primary comammox culture (Nitrospira inopinata), new comammox enrichments, and cultivation-independent approaches in collaboration with the group of Michael Wagner.
Selected publications on this topic:
Daims H, Lebedeva EV, Pjevac P, Han P, Herbold C, Albertsen M, Jehmlich N, Palatinszky M, Vierheilig J, Bulaev A, Kirkegaard RH, von Bergen M, Rattei T, Bendinger B, Nielsen PH, Wagner M. 2015. Complete nitrification by Nitrospira bacteria. Nature 528: 504-509.
Kits KD, Sedlacek CJ, Lebedeva EV, Han P, Bulaev A, Pjevac P, Daebeler A, Romano S, Albertsen M, Stein LY, Daims H, Wagner M. 2017. Kinetic analysis of a complete nitrifier reveals an oligotrophic lifestyle. Nature 549: 269-272.
Kits KD, Jung M-Y, Vierheilig J, Pjevac P, Sedlacek CJ, Liu S, Herbold CW, Stein LY, Richter A, Wissel H, Brüggemann N, Wagner M, Daims H. 2019. Low yield and abiotic origin of N2O formed by the complete nitrifier Nitrospira inopinata. Nat. Commun. 10:1836.
Pjevac P, Schauberger C, Poghosyan L, Herbold CW, van Kessel MAHJ, Daebeler A, Steinberger M, Jetten MSM, Lücker S, Wagner M, Daims H. 2017. AmoA-targeted polymerase chain reaction primers for the specific detection and quantification of comammox Nitrospira in the environment. Front. Microbiol. 8:1508.
 Ecophysiology of NOB. The elusive and still barely studied NOB are the Big Unknown of the N cycle. We unravel their metabolism by analyzing (meta)genomes and by testing genome-based hypotheses in wet lab experiments. Traditionally, NOB were perceived as highly specialized and metabolically restricted microorganisms. In contrast, we have found unexpected physiological features that far exceed nitrification and revealed previously overlooked ecological functions of NOB. For example, Nitrospira moscoviensis can grow on H2 as sole energy source and electron donor by catalyzing the oxyhydrogen (“Knallgas”) reaction (Koch et al. 2014, Science). This organism also turned out to be extremely versatile, being able to concomitantly oxidize nitrite and formate and to simultaneously use oxygen and nitrate as terminal electron acceptors (Koch et al. 2015, PNAS).
Ecophysiology of NOB. The elusive and still barely studied NOB are the Big Unknown of the N cycle. We unravel their metabolism by analyzing (meta)genomes and by testing genome-based hypotheses in wet lab experiments. Traditionally, NOB were perceived as highly specialized and metabolically restricted microorganisms. In contrast, we have found unexpected physiological features that far exceed nitrification and revealed previously overlooked ecological functions of NOB. For example, Nitrospira moscoviensis can grow on H2 as sole energy source and electron donor by catalyzing the oxyhydrogen (“Knallgas”) reaction (Koch et al. 2014, Science). This organism also turned out to be extremely versatile, being able to concomitantly oxidize nitrite and formate and to simultaneously use oxygen and nitrate as terminal electron acceptors (Koch et al. 2015, PNAS).
 Microbe-microbe interactions. In natural and engineered ecosystems, NOB never occur alone but are parts of complex microbial communities including also ammonia oxidizers and other (e.g., heterotrophic) microorganisms. Thus, an important goal of our research is to unravel possible biological interactions between NOB and these other microbial players.
Microbe-microbe interactions. In natural and engineered ecosystems, NOB never occur alone but are parts of complex microbial communities including also ammonia oxidizers and other (e.g., heterotrophic) microorganisms. Thus, an important goal of our research is to unravel possible biological interactions between NOB and these other microbial players.
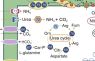
 Recently we found that the genomes of several Nitrospira and other NOB encode ureases and urea transporters, which enable these NOB to convert urea to ammonia and CO2. In co-culture experiments we showed that Nitrospira possessing urease can provide urease-negative ammonia oxidizers with ammonia. This leads to a reciprocal feeding interaction that allows the partners to grow by nitrifying urea to nitrate. Interestingly, the NOB start nitrification in this previously unknown type of interaction between nitrifiers (Koch et al. 2015, PNAS). Intriguingly, reciprocal feeding between NOB and AOB was also observed with cyanate, an environmentally widespread but previously overlooked substrate for nitrification (Palatinszky et al. 2015, Nature).
Recently we found that the genomes of several Nitrospira and other NOB encode ureases and urea transporters, which enable these NOB to convert urea to ammonia and CO2. In co-culture experiments we showed that Nitrospira possessing urease can provide urease-negative ammonia oxidizers with ammonia. This leads to a reciprocal feeding interaction that allows the partners to grow by nitrifying urea to nitrate. Interestingly, the NOB start nitrification in this previously unknown type of interaction between nitrifiers (Koch et al. 2015, PNAS). Intriguingly, reciprocal feeding between NOB and AOB was also observed with cyanate, an environmentally widespread but previously overlooked substrate for nitrification (Palatinszky et al. 2015, Nature).
 To date, little is known about the interactions of nitrifiers and heterotrophic microorganisms, which might be commensalistic, mutualistic, or even parasitic. In activated sludge, we identified a novel alphaproteobacterial predator related to Micavibrio, which specifically attacks the microcolonies of NOB from Nitrospira lineage I (related to Nitrospira defluvii) and apparently feeds on them. This first known example of a microbial predator hunting for nitrifiers suggests that nitrification is not only affected by environmental conditions, but also is subject to biological control through predator-prey interactions (Dolinsek et al. 2013, Appl. Environ. Microbiol.).
To date, little is known about the interactions of nitrifiers and heterotrophic microorganisms, which might be commensalistic, mutualistic, or even parasitic. In activated sludge, we identified a novel alphaproteobacterial predator related to Micavibrio, which specifically attacks the microcolonies of NOB from Nitrospira lineage I (related to Nitrospira defluvii) and apparently feeds on them. This first known example of a microbial predator hunting for nitrifiers suggests that nitrification is not only affected by environmental conditions, but also is subject to biological control through predator-prey interactions (Dolinsek et al. 2013, Appl. Environ. Microbiol.).
Selected publications on these topics:
Kitzinger K, Koch H, Lücker S, Sedlacek CJ, Herbold C, Schwarz J, Daebeler A, Mueller AJ, Lukumbuzya M, Romano S, Leisch N, Karst SM, Kirkegaard R, Albertsen M, Nielsen PH, Wagner M, Daims H. 2018. Characterization of the first “Candidatus Nitrotoga” isolate reveals metabolic versatility and separate evolution of widespread nitrite-oxidizing bacteria. mBio 9: e01186-18.
Koch H, Lücker S, Albertsen M, Kitzinger K, Herbold C, Spieck E, Nielsen PH, Wagner M, Daims H. 2015. Expanded metabolic versatility of ubiquitous nitrite-oxidizing bacteria from the genus Nitrospira. Proc. Natl. Acad. Sci. USA 112: 11371-11376.
Palatinszky M, Herbold C, Jehmlich N, Pogoda M, Han P, von Bergen M, Lagkouvardos I, Karst SM, Galushko A, Koch H, Berry D, Daims H, Wagner M. 2015. Cyanate as an energy source for nitrifiers. Nature 524: 105-108.
Koch H, Galushko A, Albertsen M, Schintlmeister A, Gruber-Dorninger C, Lücker S, Pelletier E, Le Paslier D, Spieck E, Richter A, Nielsen PH, Wagner M, Daims H. 2014. Growth of nitrite-oxidizing bacteria by aerobic hydrogen oxidation. Science 345: 1052-1054.
Dolinsek J, Lagkouvardos I, Wanek W, Wagner M, Daims H. 2013. Interactions of nitrifying bacteria and heterotrophs: Identification of a Micavibrio-like, putative predator of Nitrospira. Appl. Environ. Microbiol. 79: 2027-2037.
 Evolution. By comparative genome analyses we illuminate the evolutionary history of NOB, which is unexpectedly complex and interwoven with other N cycle processes such as aerobic ammonia oxidation (Daims et al. 2015, Nature) and anammox (Lücker et al. 2010, PNAS). Given the large phylogenetic diversity of known NOB with six genera from four bacterial phyla, the evolution of nitrite oxidation is a prime example of the modularity of a major biogeochemical cycle. For example, the key enzyme for nitrite oxidation, nitrite oxidoreductase (NXR), evolved multiple times independently (Lücker et al. 2010, PNAS). Nevertheless, similar types of NXR occur in phylogenetically distant NOB, suggesting that lateral gene transfer of these “nitrite oxidation modules” played a significant role in the evolution of NOB (Lücker et al. 2013, Front. Microbiol.; Sorokin et al. 2012, ISME J.).
Evolution. By comparative genome analyses we illuminate the evolutionary history of NOB, which is unexpectedly complex and interwoven with other N cycle processes such as aerobic ammonia oxidation (Daims et al. 2015, Nature) and anammox (Lücker et al. 2010, PNAS). Given the large phylogenetic diversity of known NOB with six genera from four bacterial phyla, the evolution of nitrite oxidation is a prime example of the modularity of a major biogeochemical cycle. For example, the key enzyme for nitrite oxidation, nitrite oxidoreductase (NXR), evolved multiple times independently (Lücker et al. 2010, PNAS). Nevertheless, similar types of NXR occur in phylogenetically distant NOB, suggesting that lateral gene transfer of these “nitrite oxidation modules” played a significant role in the evolution of NOB (Lücker et al. 2013, Front. Microbiol.; Sorokin et al. 2012, ISME J.).
Selected publications on this topic:
Lücker S, Nowka B, Rattei T, Spieck E, Daims H. 2013. The genome of Nitrospina gracilis illuminates the metabolism and evolution of the major marine nitrite oxidizer. Front. Microbiol. 4: 27.
Sorokin DY, Lücker S, Vejmelkova D, Kostrikina NA, Kleerebezem R, Rijpstra WI, Damsté JS, Le Paslier D, Muyzer G, Wagner M, van Loosdrecht MC, Daims H. 2012. Nitrification expanded: Discovery, physiology, and genomics of a nitrite-oxidizing bacterium from the phylum Chloroflexi. ISME J. 6: 2245-2256.
Lücker S, Wagner M, Maixner F, Pelletier E, Koch H, Vacherie B, Rattei T, Sinninghe Damsté JS, Spieck E, Le Paslier D, Daims H. 2010. A Nitrospira metagenome illuminates the physiology and evolution of globally important nitrite-oxidizing bacteria. Proc. Natl. Acad. Sci. USA 107: 13479-13484.
 Diversity and ecological niche partitioning. NOB are a diverse functional group, and recently we identified new lineages of NOB or revealed their functional importance in wastewater treatment (Sorokin et al. 2012, ISME J.; Lücker et al. 2015, ISME J.). We also discovered a huge diversity of closely related coexisting NOB (Nitrospira) in natural habitats such as soils (Pester et al. 2014, Environ. Microbiol.) and in wastewater treatment plants, where we showed distinct ecophysiological features of the diverse Nitrospira populations (Gruber-Dorninger et al. 2015, ISME J.). To analyze the microdiversity of these uncultured NOB in detail, we identify NOB microcolonies in situ in activated sludge by an innovative approach based on isotope labeling and Raman microspectroscopy. The identified NOB microcolonies are extracted by micromanipulation and their metagenomes are sequenced by single-cell genomics techniques. The obtained genomic data are analyzed with focus on NOB population genetics and biological mechanisms that support such complex and stable communities of coexisting NOB. Since closely attached microbial symbionts are co-extracted with the NOB microcolonies, their genomes are sequenced together with the NOB and allow novel insights into the biological interactions between NOB and other microorganisms. This research is carried out in close collaboration with Michael Wagner at DOME and Tanja Woyke at JGI (Walnut Creek, USA).
Diversity and ecological niche partitioning. NOB are a diverse functional group, and recently we identified new lineages of NOB or revealed their functional importance in wastewater treatment (Sorokin et al. 2012, ISME J.; Lücker et al. 2015, ISME J.). We also discovered a huge diversity of closely related coexisting NOB (Nitrospira) in natural habitats such as soils (Pester et al. 2014, Environ. Microbiol.) and in wastewater treatment plants, where we showed distinct ecophysiological features of the diverse Nitrospira populations (Gruber-Dorninger et al. 2015, ISME J.). To analyze the microdiversity of these uncultured NOB in detail, we identify NOB microcolonies in situ in activated sludge by an innovative approach based on isotope labeling and Raman microspectroscopy. The identified NOB microcolonies are extracted by micromanipulation and their metagenomes are sequenced by single-cell genomics techniques. The obtained genomic data are analyzed with focus on NOB population genetics and biological mechanisms that support such complex and stable communities of coexisting NOB. Since closely attached microbial symbionts are co-extracted with the NOB microcolonies, their genomes are sequenced together with the NOB and allow novel insights into the biological interactions between NOB and other microorganisms. This research is carried out in close collaboration with Michael Wagner at DOME and Tanja Woyke at JGI (Walnut Creek, USA).
Selected publications on this topic:
Gruber-Dorninger C, Pester M, Kitzinger K, Savio DF, Loy A, Rattei T, Wagner M, Daims H. 2015. Functionally relevant diversity of closely related Nitrospira in activated sludge. ISME J. 9: 643-655.
Lücker S, Schwarz J, Gruber-Dorninger C, Spieck E, Wagner M, Daims H. 2015. Nitrotoga-like bacteria are previously unrecognized key nitrite oxidizers in full-scale wastewater treatment plants. ISME J. 9: 708-720.
Pester M, Maixner F, Berry D, Rattei T, Koch H, Lücker S, Nowka B, Richter A, Spieck E, Lebedeva E, Loy A, Wagner M, Daims H. 2014. NxrB encoding the beta subunit of nitrite oxidoreductase as functional and phylogenetic marker for nitrite-oxidizing Nitrospira. Environ. Microbiol., 16: 3055-3071.
Sorokin DY, Lücker S, Vejmelkova D, Kostrikina NA, Kleerebezem R, Rijpstra WI, Damsté JS, Le Paslier D, Muyzer G, Wagner M, van Loosdrecht MC, Daims H. 2012. Nitrification expanded: Discovery, physiology, and genomics of a nitrite-oxidizing bacterium from the phylum Chloroflexi. ISME J. 6: 2245-2256.
Other selected publications on nitrifying microorganisms:
Mussmann M, Brito I, Pitcher A, Sinninghe Damsté JS, Hatzenpichler R, Richter A, Nielsen JL, Nielsen PH, Müller A, Daims H, Wagner M, Head IM. 2011. Thaumarchaeotes abundant in refinery nitrifying sludges express amoA but are not obligate autotrophic ammonia oxidizers. Proc. Natl. Acad. Sci. USA 108: 16771-16776.
Hatzenpichler R, Lebedeva EV, Spieck E, Stoecker K, Richter A, Daims H, Wagner M. 2008. A moderately thermophilic ammonia-oxidizing crenarchaeote from a hot spring. Proc. Natl. Acad. Sci. USA 105: 2134-2139.
Maixner F, Wagner M, Lücker S, Pelletier E, Schmitz-Esser S, Hace K, Spieck E, Konrat R, Le Paslier D, Daims H. 2008. Environmental genomics reveals a functional chlorite dismutase in the nitrite-oxidizing bacterium "Candidatus Nitrospira defluvii". Environ. Microbiol. 10: 3043-3056.
Mlynek G, Sjöblom B, Kostan J, Füreder S, Maixner F, Gysel K, Furtmüller PG, Obinger C, Wagner M, Daims H, Djinovic-Carugo K. 2011. Unexpected diversity of chlorite dismutases: A catalytically efficient dimeric enzyme from Nitrobacter winogradskyi. J. Bacteriol. 193: 2408-2417.
Microscopy and image analysis methods for characterizing microbial communities
 Fluorescence labeling methods are essential tools in modern microbial ecology and medical microbiology. A particularly well-known example is fluorescence in situ hybridization (FISH) with rRNA-targeted probes for the cultivation-independent identification, detection, and quantification of microorganisms. To unleash the full power of these labeling techniques, one must extract quantitative data from the resulting fluorescence micrographs. Common applications are measuring the brightness of the single-cell fluorescence labeling or quantifying the in situ abundances of microbial populations. More advanced possibilities arise if fluorescence labeling is combined with high-quality confocal laser scanning microscopy, which enables three-dimensional analyses of biofilms and other spatially complex samples.
Fluorescence labeling methods are essential tools in modern microbial ecology and medical microbiology. A particularly well-known example is fluorescence in situ hybridization (FISH) with rRNA-targeted probes for the cultivation-independent identification, detection, and quantification of microorganisms. To unleash the full power of these labeling techniques, one must extract quantitative data from the resulting fluorescence micrographs. Common applications are measuring the brightness of the single-cell fluorescence labeling or quantifying the in situ abundances of microbial populations. More advanced possibilities arise if fluorescence labeling is combined with high-quality confocal laser scanning microscopy, which enables three-dimensional analyses of biofilms and other spatially complex samples.
 We develop a set of digital image analysis algorithms for analyzing fluorescence-labeled microorganisms, which are bundled in our image analysis software daime. In combination with suitable protocols for epifluorescence and confocal microscopy, these tools enable the precise quantification of key features of microbial populations and communities. Notably, we have developed image analysis routines for quantifying the spatial arrangement patterns of microorganisms in biofilms, which often are hallmarks of important biological interactions such as mutualistic symbioses or competition for resources. Our software also allows the interactive 3D visualization of confocal image stacks and integrates this feature seamlessly with 3D image analysis functionality. Further details and the download links are available on the daime website.
We develop a set of digital image analysis algorithms for analyzing fluorescence-labeled microorganisms, which are bundled in our image analysis software daime. In combination with suitable protocols for epifluorescence and confocal microscopy, these tools enable the precise quantification of key features of microbial populations and communities. Notably, we have developed image analysis routines for quantifying the spatial arrangement patterns of microorganisms in biofilms, which often are hallmarks of important biological interactions such as mutualistic symbioses or competition for resources. Our software also allows the interactive 3D visualization of confocal image stacks and integrates this feature seamlessly with 3D image analysis functionality. Further details and the download links are available on the daime website.
Selected publications on this topic:
Riva A, Kuzyk O, Forsberg E, Siuzdak G, Pfann C, Herbold C, Daims H, Loy A, Warth B, Berry D. 2019. A fiber-deprived diet disturbs the fine-scale spatial architecture of the murine colon microbiome. Nat. Commun. 10: 4366.
Remus-Emsermann MNP, Lücker S, Müller DB, Potthoff E, Daims H, Vorholt JA. 2014. Spatial distribution analyses of natural phyllosphere-colonizing bacteria on Arabidopsis thaliana revealed by fluorescence in situ hybridization. Environ. Microbiol. 16: 2329-2340.
Almstrand R, Daims H, Persson F, Sörensson F, Hermansson M. 2013. New methods for analysis of spatial distribution and co-aggregation of microbial populations in complex biofilms. Appl. Environ. Microbiol. 79: 5978-5987.
Schillinger C, Lux R, Riep B, Kikhney J, Petrich A, Friedmann A, Wolinsky LE, Gobel UB, Daims H, Moter A. 2012. Co-localized or randomly distributed? Pair cross correlation of in vivo grown subgingival biofilm bacteria quantified by digital image analysis. PLoS One 7: e37583.
Daims H, Wagner M. 2011. In situ techniques and digital image analysis methods for quantifying spatial localization patterns of nitrifiers and other microorganisms in biofilm and flocs. Methods Enzymol. 496: 185-215.
Daims H, Lücker S, Wagner M. 2006. daime, a novel image analysis program for microbial ecology and biofilm research. Environ. Microbiol. 8: 200-213.
Current research projects
The Comammox Research Platform
Illuminating the ecology of nitrite-oxidizing bacteria in soil and aquatic ecosystems
Joining the team
Information on open research positions can be found here. If you are interested in joining our team with your own fellowship, please check out our PhD & postdoc program and get in touch with Holger for details.